La photosynthèse
2. La photosynthèse
2.1 – définition de la photosynthèse :
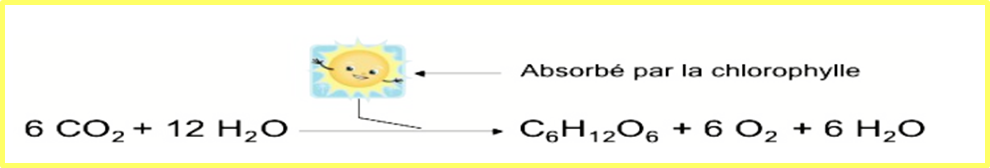
C’est le processus bioénergétique qui permet aux plantes, aux algues et à certaines bactéries de synthétiser de la matière organique grâce à lumière. - La plante a besoin du CO2 de l’air, de l’eau et les minéraux du sol. – Les végétaux sont autotrophes pour le carbone, ce qui permet la libération de molécules d’O2.
La photosynthèse comprend l’utilisation de l’énergie lumineuse pour la réduction du dioxyde de carbone par l’eau, donneur d’hydrogène (ou d’électrons + protons), avec synthèse de glucides et libération d’oxygène. Cette réaction est réalisée dans des organites intracellulaires spécialisés, les chloroplastes, porteurs de pigments photorécepteurs, tels que les chlorophylles, les caroténoïdes.
La photosynthèse se déroule en deux phases : lors de la première, les réactions dépendantes de la lumière captent l'énergie lumineuse et l'utilisent pour produire une coenzyme réductrice, le NADPH, et une coenzyme qui stocke l'énergie chimique, l'ATP, tandis que, lors de la seconde phase, les réactions indépendantes de la lumière utilisent ces coenzymes pour absorber et réduire le dioxyde de carbone.
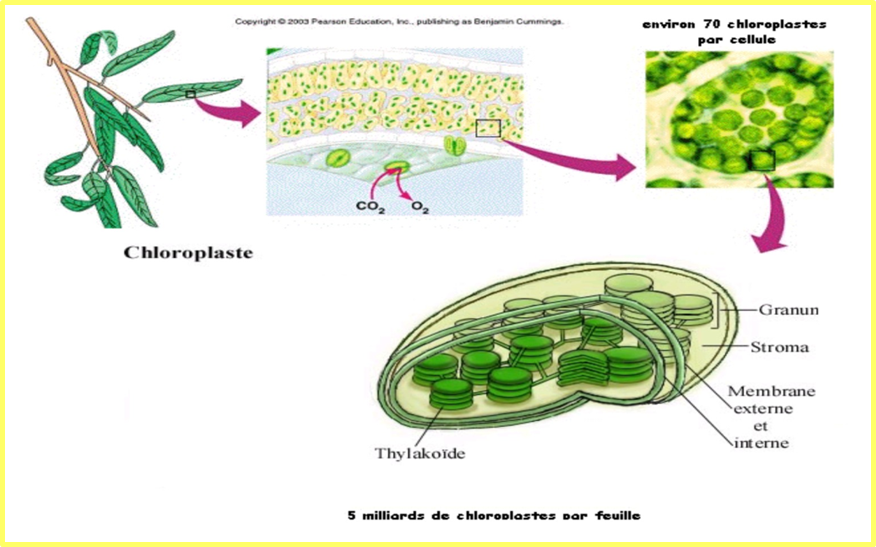
2.2 Localisation de la photosynthèse
Dans les chloroplastes des cellules, il y a des structures membranaires, les thylakoïdes qui sont très riches en pigments et en protéines.
La nervure médiane, très en relief comme chez beaucoup de dicotylédones contient principalement des tissus conducteurs de la sève brute (xylème) et de la sève élaborée (phloème).
Ces tissus sont protégés par des tissus de soutien. De part et d'autre de cette nervure, le limbe est formé par du parenchyme palissadique (face supérieure) et du parenchyme lacuneux (face inférieure). La surface de la feuille est recouverte d'une cuticule cireuse et imperméable qui protège la feuille de l'excès d'évaporation d'eau et réduit l'absorption de la lumière bleue et ultraviolette afin de limiter l'échauffement de la plante.
L'épiderme transparent permet à la lumière de le traverser pour mieux atteindre le mésophylle, où se déroule l'essentiel de la photosytnhèse. Les échanges de gaz sont assurés par les stomates.
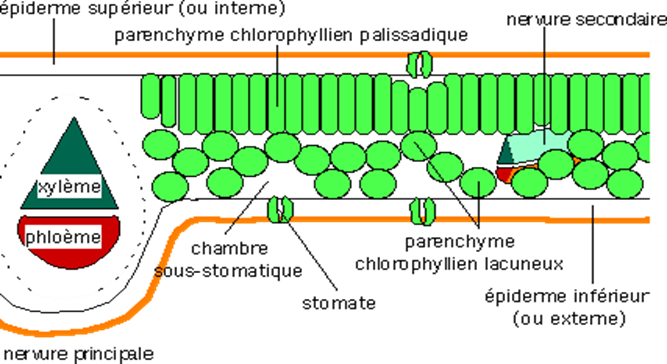
Structure schématique d'une feuille de Dicotylédones
La feuille de Monocotylédones est le plus souvent
caractérisée par la présence de nervures parallèles et de taille sensiblement
équivalente. La feuille étant souvent verticale (Graminées, Liliacées, etc.) il
n'y a pas de distinction nette entre les deux sortes de parenchymes
chlorophylliens
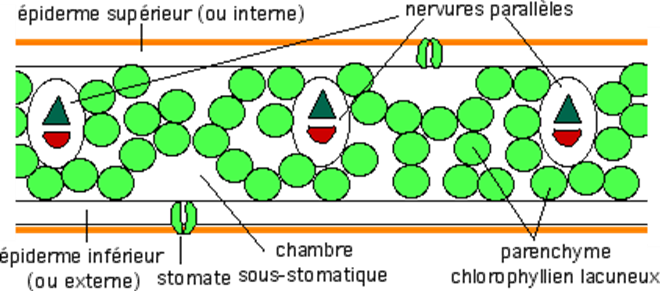
Structure schématique d'une feuille de Monocotylédones.
2.2.1 Le parenchyme chlorophyllien
Chez la plupart des Dicotylédones (feuilles en position plus ou moins horizontales), il existe un parenchyme chlorophyllien palissadique à la face supérieure et un parenchyme lacuneux à la face inférieure. Chez beaucoup de Monocotylédones (feuilles en position verticale) on n'observe le plus souvent qu'un seul parenchyme, plus ou moins lacuneux. Le parenchyme chlorophyllien, lieu de la photosynthèse, est mis en relation dans la feuille vers l'extérieur (échanges gazeux) par les stomates et vers l'intérieur (apport d'eau et de sels minéraux et transport des assimilât) par les nervures
2.2.2 Organites et membranes de la photosynthèse
- Chloroplastes et thylakoïdes
La photosynthèse se déroule dans des organites appelés chloroplastes. Une cellule typique de plante contient environ dix à cent chloroplastes. Ces derniers sont enveloppés par une membrane interne et une membrane externe de nature phospholipidique et séparées par un espace intermembranaire. L'intérieur du chloroplaste est constitué d'un fluide aqueux appelé stroma. Dans le stroma se trouvent des thylakoïdes empilés en grana où siège la photosynthèse. Les thylakoïdes sont en forme de disques aplatis délimités par une membrane contenant l'espace thylakoïde ou lumen. La photosynthèse se déroule précisément dans la membrane des thylakoïdes, qui contient des complexes protéiques membranaires intégraux et périphériques ainsi que les pigments qui absorbent l'énergie lumineuse et constituent les photosystèmes.
Le chloroplaste est le siège de la photosynthèse. Il absorbe l’énergie lumineuse pour fixer le carbone inorganique (CO2) sous forme de glucose, au cours de ce processus de l'énergie chimique sous forme d'adénosine triphosphate (ATP) est également produite. Cet ATP intervient dans la phase photochimique de la photosynthèse.
Le chloroplaste absorbe l'ensemble du spectre de la lumière visible mis à part le vert, raison pour laquelle les feuilles des plantes ont un aspect vert. La chlorophylle se trouve dans la membrane des thylakoïdes. Les différentes étapes de la photosynthèse qui convertissent la lumière en énergie chimique se déroulent dans les thylakoïdes tandis que les étapes de conversion de l'énergie en glucide se déroulent dans le stroma du chloroplaste.
- Les pigments assimilateurs
Les pigments photosynthétiques ou pigments assimilateurs, sont les composés chimiques permettant la transformation de l'énergie lumineuse en énergie chimique chez les organismes effectuant la photosynthèse. Lorsqu'un photon heurte une molécule de pigment photosynthétique, son énergie excite un atome de cette molécule, et la fait passer à un état excité, de niveau énergétique élevé. L'énergie accumulée dans la molécule de pigment est libérée lors du retour à l'état fondamental de la molécule, il y a trois voies de libération de cette énergie en plus d'un dégagement de chaleur.
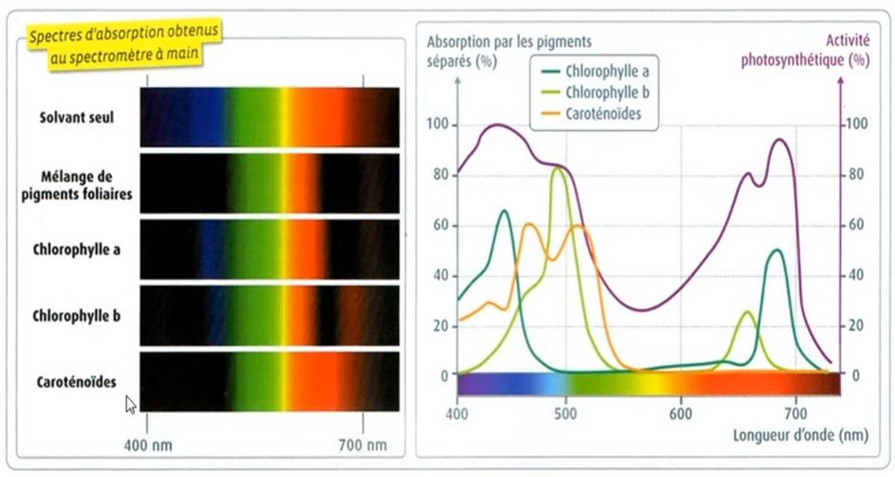
Les principaux pigments présents chez les végétaux sont les chlorophylles (a et b) et les caroténoïdes. En fonction de ces propriétés moléculaires, chaque pigment absorbera des photons ayant des longueurs d’ondes différentes, (entre 300 et 800 nm) ce qui permet de définir le spectre d’absorption de chaque pigment.
- Les pigments chlorophylliens
La chlorophylle de couleur verte, est disposée régulièrement dans les différents saccules des granas (thylacoïdes) des chloroplastes. Elles sont liées aux protéines membranaires des thylakoides. Elle consiste en plusieurs molécules encastrées en partie dans des couches de lipides qui les séparent.
Il existe quatre types de chlorophylle, qui sont bien souvent mélangés dans les granas. La plus commune et la plus importante dans le processus est la chlorophylle a. Elle est d'un vert profond (C55H72O5N4Mg). La chlorophylle b (C55H70O6N4Mg) quant à elle, est un pigment jaune-vert. Les plantes à graines, par exemple, contiennent trois parties de chlorophylle a pour une de chlorophylle b. Pour ce qui est des chlorophylles c et d, on les retrouve respectivement dans les algues brunes et algues rouges, mélangées à de la chlorophylle a.
.
Diverses radiations composent la lumière blanche et chacune a une action
spécifique. Leur énergie est inversement proportionnelle à leur longueur
d'onde, ainsi, le spectre rouge
contient moins d'énergie
que le bleu. Les radiations rouges
(600nm) et indigo (400-450nm) sont absorbées par la chlorophylle et sont les plus efficaces. Les vertes n'ont aucun effet,
car elles sont soit réfléchies, soit elles passent
directement au travers de la plante.
Finalement, de toute cette lumière,
20% est réfléchie, 10% transmise
et 70% est absorbée (20% dissipée en chaleur, 48% en fluorescence et 2% seulement à la photosynthèse)
2.3 Processus de la photosynthèse
La photosynthèse se déroule en deux phases :
* une phase photochimique dite « lumineuse ou claire » dépendante de la lumière et se déroule dans les membranes des thylakoïdes. Cette phase permet la photolyse de l’eau, avec dégagement d’oxygène, synthèse d’ATP et réduction de NADP+ en NADPH + H+ ;
* Une phase chimique, dite « obscure », indépendante de la lumière, le cycle de Calvin utilise l'énergie chimique produite dans la première phase pour synthétiser des glucides à partir du CO2 et se déroule dans le stroma des chloroplastes
2.3.1- La phase lumineuse
Cette phase a besoin de lumière et de la chlorophylle a pour s'effectuer. Les molécules de pigment sont organisées en complexes dans la membrane du thylakoide. On trouve deux types principaux de complexes, les photosystèmes I et II (PSI et PSII). Chaque complexe est organisé autour d’une paire de molécule de chlorophylle a particulière, située dans le centre réactionnel du photosystème. Le photosystème II contenant de la chlorophylle P680 (absorbant mieux à 680 nm) et le photosystème I contenant de la chlorophylle P700.
Photosystème (PS) : C’est l’association entre une chlorophylle avec des protéines et d'autres molécules organiques. Elle comprend une antenne collectrice d’énergie lumineuse et un centre réactionnel. Les deux photosystèmes PSII et PSI interviennent successivement dans la phase photochimique.
L’antenne collectrice est constituée de protéines particulières ayant la propriété d’établir des liaisons stables avec les pigments chlorophylliens. Elle assure la capture efficace des photons et transmet l’énergie d’excitation au centre réactionnel.
Les différents pigments peuvent recevoir des photons de longueur d’onde différente ; compte tenu de leur proximité, une molécule excitée peut transmettre son excitation à des molécules voisines par résonances. Ce transfert se fera spontanément depuis un pigment absorbant les basses longueurs d’ondes (les plus énergétiques) vers les pigments absorbant les hautes longueurs d’ondes. L’antenne collectrice permet donc de faire converger l’énergie lumineuse vers les chlorophylles a du centre réactionnel
Les antennes, incluses dans la membrane, constituent les complexes collecteurs de lumière (Light harvesting complex, LHC). LHC I et LHC II sont ainsi associées respectivement au PSI et PSII.
Le centre réactionnel est constitué par un dimère de chlorophylle a, auxquelles sont adjointes un certain nombre de protéines assurant des fonctions diverses : transfert d’électrons, fonctions de régulation… etc. Cet ensemble forme le cœur du photosystème.
Le photosystème II
La structure de ce photosystème est complexe, comprenant une vingtaine de protéines. L‘antenne du PSII est constituée de deux parties :
* l’une dite antenne interne ou proximale, est étroitement liée au cœur du PSII. Elle renferme des protéines assez semblables, CP43 et CP47, liées a 40-50 molécules de Chl a et à une dizaine de molécules de B-carotène (la chlorophylle b est absente)
* l’autre dite antenne périphérique ou distale, constitue le LHC II au sens strict. Elle est de taille beaucoup plus grande et renferme la moitié de la Chl et le tiers des protéines des Thylacoides (protéines liant la Chl a et la Chl b ou binding proteins CAB).
Le pigment actif du centre réactionnel du PSII, P680 est un dimère de chlorophylle a. Il constitue le donneur primaire d’électrons. Son potentiel d’oxydoréduction très élevé lui permet de recevoir les électrons issus de l’eau. 2H2O O2 + 4H+ + 4e-
Le complexe Z ou complexe d’émission de l’oxygène (OEC, Oxygen Evolving Complex), se trouve dans le lumen où il libère les protons issus de la scission de l’eau. Les électrons sont transmis du complexe Z à P680 par l’intermédiaire d’une Tyrosine (Tyrz ou Yz) appartenant à la protéine D1.
L’accepteur primaire est une molécule de Phéophytine a. de là, l’électron est successivement transféré sur deux plastoquinones, nommées QA et QB, respectivement liées à deux protéines D2 et D1. Le rôle des deux quinones QA et QB est de stoquer les électrons, reçus un à un et de les transmettre par paires sur l’accepteur suivant, la plastoquinone.
La plastoquinone est une coenzyme de la famille des quinones participant à la chaîne de transport d'électrons de la photophosphorylation au cours de la photosynthèse. Elle est localisée dans la membrane des thylakoïdes des chloroplastes. Elle reçoit les électrons provenant du photosystème II et les transfère sur le complexe cytochrome b6f.
Entre PSII et PSI : le complexe b6f
Le complexe b6f (le complexe cytochrome b6f) est un complexe enzymatique (oxydoréductase) qui intervient dans le transfert des électrons entre deux centres réactionnels, du photosystème II au
photosystème I tout en pompant des protons à travers la membrane des thylakoïdes depuis le stroma des chloroplastes vers le lumen des thylakoïdes. C'est la circulation des électrons à travers le complexe cytochrome b6f qui génère le gradient de concentration en protons, et donc le gradient électrochimique, à travers la membrane des thylakoïdes.
Il est constitué de quatre éléments au moins :
* Une protéine Fe-S qui renferme un centre 2Fe-2S, c’est elle qui reçoit les électrons provenant de la plastoquinone. Ils jouent le rôle de « condensateurs » biologiques, capables de se charger et de se décharger en passant d'un état d'oxydation à un autre au cours de réactions d'oxydoréduction.
* Le cytochrome f : reçoit les électrons de la protéine Fe-S et les transmets à la photocyanine qui les véhiculera jusqu’au PS I.
* le cytochrome b6 : il se trouve sous deux formes qui participent au transport cyclique d’électrons en recevant des électrons provenant de la ferédoxine réduite et au cycle Q.
Le cycle Q : la réduction de PQ (Plastoquinones) en PQH2 (Plastoquinols) et sa réoxydation en plastoquinones (PQ) (l’oxydation se fait par la b6f), accompagnées de capture et de libération de H+, forment un cycle, le cycle Q. Les H+ (prélevés dans le stroma) sont émis dans le lumen, et le cycle Q fonctionne comme une véritable pompe à protons.
La plastocyanine : c’est une petite protéine renfermant un atome de Cu. Très peu liée à la membrane du thylacoide et pratiquement en solution dans le lumen, elle transfert les électrons du complexe b6f au PS I.
Le photosystème I
Il est composé d’une vingtaine de polypeptides distribués entre un centre réactionnel (P700) et une antenne (LHC I).
Après interception d’un photon, P700 (donneur primaire) est oxydé en P700+ , ce qui lui permet d’être réduit par la plastocyanine (donneur secondaire). De la l’électron est transféré sur un accepteur secondaire, la phylloquinone (VitamineK1) ; puis il passe sur la protéine Fx. Le trajet de l’électron passe par deux autres protéines FB et FA avant d’atteindre la ferrédoxine.
La ferrédoxine : c’est un transporteur hydrophile très mobile, qui sert à distribuer le pouvoir réducteur dans la cellule. Par l’intermédiaire de l’enzyme ferrédoxine-NADP+ réductase (FNR) elle va réduire le NADP+ pour donner le NADPH (libéré dans le stroma) qui représente le produit final de la chaine de transport d’électrons.
Elle peut aussi céder ses électrons au complexe b6f engendrant ainsi un transport d’électron cyclique. Les électrons retourneront sur P700 par l’intermédiaire de la plastocyanine.
La photophosphorylation
La synthèse d'ATP dans le chloroplaste s'effectue au niveau du complexe ATP synthase associée à la membrane du thylakoïde. Cette synthèse d'ATP dans le chloroplaste est appelée photophosphorylation. Il existe deux types de phosphorylations :
a/ La phosphorylation acyclique est associée au trajet linéaire des électrons entre H2O et NADP+ au cours duquel, par molécule de H2O détruite, 4e sont transférés et 2 NADPH formés. A ce transfert est associé la synthèse d’un certain nombre (n) de molécules d’ATP.
b/La phosphorylation cyclique : elle ne fait intervenir que le photosystème I, le complexe cytochrome b6f et la plastocyanine ; le photosystème I absorbe l'énergie lumineuse, ce qui permet d'exciter un électron, transféré à un complexe cytochrome b6f auquel il cède son énergie en générant un gadient électrochimique utilisé par une ATP synthase pour produire de l'ATP, avant de revenir au photosystème I via une plastocyanine. Ces réactions ne produisent ni O2 ni NADPH.
2.3.2- Phase obscure
C’est une série de réactions biochimiques prenant place dans le stroma des chloroplastes des organismes photosynthétiques. Durant la photosynthèse, l’énergie de la lumière a été convertie en énergie chimique conservée dans l’ATP et le NADPH. Le cycle de Calvin, indépendant de la lumière, utilise l’énergie de ces transporteurs pour transformer le dioxyde de carbone en composés organiques qui peuvent être utilisés par l’organisme.
Au cours de cette phase obscure, la ribulose biphosphate s’additionne au dioxyde de carbone gazeux (CO2) présent dans l’air pour produire des composés organiques, principalement des hydrates de carbone ou des sucres
a) Fixation du dioxyde de carbone
Cette réaction est catalysée par la RubisCO (Ribulose bis Phosphate Carboxylase/Oxygénase). Cette enzyme a deux fonctions. C'est la fonction "carboxylase" qui nous intéresse ici. La fonction "oxygénase" joue un rôle important dans la photorespiration.
La RubisCO incorpore une molécule de CO2 dans un composé en C5, le ribulose bis phosphate (RUBP) pour donner deux molécules d'un composé en C3, l'acide phosphoglycérique (APG), après passage par un intermédiaire réactionnel en C6.
b) La réduction de l'APG en trioses-P
L'APG, le premier composé formé par l'incorporation du CO2, n'est pas un sucre. Une étape importante consiste donc en la réduction de l'APG. L'APG se combine avec de l'hydrogène fourni par le NADPH2, ce qui produit de l'eau et du PGAL (phosphoglycéraldéhyde ou triose phosphate). L'énergie nécessaire à ceci est fournie par l'ATP qui perd un groupe phosphate pour redevenir de l'ADP.
c) La régénération du RUBP
Le principe même du cycle de Calvin est de permettre la régénération du RUBP à partir d'une fraction des trioses formés. C'est la condition nécessaire qui permettra à nouveau de réaliser l'incorporation de nouvelles molécules de CO2.
On peut schématiser le processus de régénération du RuBP en considérant la fixation de 3 molécules de CO2 qui conduisent à la formation de 6 trioses. Sur les 6 trioses formés, 1 triose servira à la synthèse ultérieure des sucres plus complexes, les 5 autres trioses servant à régénérer 3 molécules de RUBP,
Finalement, les pentoses phosphates formés (RuP) sont convertis en RuBP grâce à l'ATP. Cette réaction de phosphorylation est catalysée par la Phosphate Ribulose Kinase (PRK).
La régénération du RuBP nécessite donc une molécule d’ATP supplémentaire par molécule de CO2 fixé.
Les molécules du PGAL contiennent des chaînes de trois atomes de carbone, donc peuvent être utilisées comme aliment par la plante, mais elle en produit bien plus qu'elle en a besoin. Le surplus est donc transformé en d'autres produits. Un des plus importants est le RDP (rubulose diphosphate) qui sert à réapprovisionner la cellule en combustible pour les prochaines réactions.
D'autres molécules de PGAL sont transformées en glucose, en fructose, en maltose, et peuvent également être liées en chaînes pour former de l'amidon ou de la cellulose. Certaines plantes se servent aussi d'une partie du PGAL pour en faire des huiles (ex. : le maïs, le lin, le ricin et l'olivier)
Les Facteurs d'influence
La photosynthèse est influencée par les facteurs de l'environnement : la lumière (source d'énergie), le CO2 (source de carbone) et la température (qui affecte l'ensemble des réactions biochimiques).
Les facteurs externes agissent indépendamment les uns des autres et le phénomène global obéit à la loi dite des "facteurs limitants" que l'on peut énoncer de la façon suivante :
Lorsqu’un processus est contrôlé par plusieurs facteurs agissants indépendamment, son intensité est limitée par le facteur qui présente la valeur minimum. Le facteur est alors limitant et la vitesse du processus est proportionnelle à la valeur de ce facteur.
L'assimilation des substances entrant en jeu lors de la photosynthèse est influencée par plusieurs facteurs.
La teneur en gaz carbonique de l'air (0,03%) est très faible, mais les végétaux sont capables de synthétiser une plus grande quantité de matière quand elle augmente. La teneur en CO2 idéale se situe aux environs de 0,1% (cette propriété est parfois utilisée dans les serres pour maximiser le rendement). Si le pourcentage est entre 2 et 5% (ou plus), il devient alors toxique et nuit au développement de la plante
Selon l'origine des plantes, l'optimum de température de leur activité photosynthétique est différent. Les plantes des régions tempérées ont un maximum qui se situe entre 15°C et 25°C, tandis que les plantes d'origine tropicale peuvent avoir un maximum qui se situe entre 30 et 45°C.
Certains végétaux ont besoin d'une lumière intense pour bien fonctionner (plantes de soleil, héliophiles) et d'autres (plantes d'ombre, sciaphiles) ont besoin de beaucoup moins de lumière et souffrent d'un excès de cette dernière.
L'approvisionnement en eau se fait par les racines pour les plantes terrestres et elle est transmise par une canalisation vers les feuilles. Un manque d'eau affecte la physiologie de la cellule et réduit le taux de photosynthèse